Felles avstamning: 'livets tre' -en mislykket hypotese
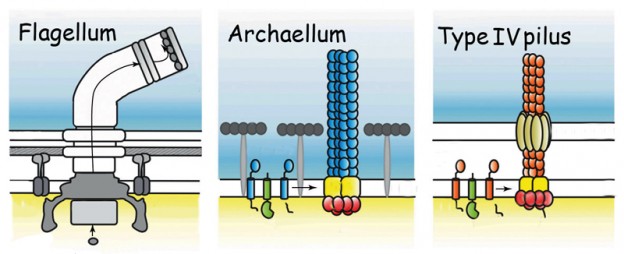 1. DNA-replikasjonsmaskineriet er ikke homologt i livets 3 domener. Bakterie celle-kjernens replisome-enzymer deler ikke en felles stamfar med de analoge komponentene i eukaryoter og archaea.
1. DNA-replikasjonsmaskineriet er ikke homologt i livets 3 domener. Bakterie celle-kjernens replisome-enzymer deler ikke en felles stamfar med de analoge komponentene i eukaryoter og archaea.
2. Bakterier og archaea skiller seg påfallende ut i kjemien til deres membranlipider. Cellemembran-fosfolipider syntetiseres av forskjellige, ikke-relaterte enzymer i bakterier og arkeaer, og gir kjemisk forskjellige membraner.
3. Sekvenser av glykolytiske enzymer er forskjellige mellom archaea, bakterier og eukaryoter. Det er ingen bevis for en felles stamfar for noen av de fire glykolytiske kinasene eller de syv enzymene som binder nukleotider.
4. Det er minst seks forskjellige autotrofiske karbonfikserings-veier. Hvis felles stamfedre holdt stikk, burde en opprinnelig Wood–Ljungdahl-sti ha vært livets eneste prinsipp for produksjon av biomasse.
5. Det er et skarpt skille i den organisatoriske kompleksiteten til cellen mellom eukaryoter, som har kompleks intracellulær inndeling, og til og med de mest sofistikerte prokaryoter (archaea og bakterier), som ikke har det.
6. En typisk eukaryot celle er omtrent 1000 ganger større i volum enn en typisk bakterie- eller arkea-celle, og fungerer under forskjellige fysiske prinsipper: fri diffusjon har liten rolle i eukaryote celler, men er avgjørende i prokaryoter.
7. Senere massiv sekvensering av mange, komplette mikrobielle genomer har avslørt nye evolusjonære fenomener, den mest grunnleggende av disse er: gjennomgripende horisontal genoverføring (HGO), i stor grad formidlet av virus og plasmider, som former genomene til archaea og bakterier og kaller på en radikal revisjon (hvis ikke oppgivelse) av Livets-Tre (LT) konseptet.
8. Forskjeller i RNA-polymerase: Prokaryoter inneholder bare tre forskjellige promoterelementer: -10, -35 promotorer og oppstrøms-elementer. Eukaryoter inneholder mange forskjellige promoter-elementer.
9. Ribosom- og ribosom-biogenese forskjeller: Selv om vi kunne identifisere E. coli-kolleger med sammenlignbar biokjemisk aktivitet for 12 gjærRibosom BiogeneseFaktorer (RBF), er det kun 2 som er kjent for å delta i bakteriell ribosom sammensetning. Dette indikerer at rekruttering av individuelle proteiner til denne veien i stor grad har vært uavhengig i de bakterielle og eukaryote linjene. (22)
I sin bok antyder Darwin at alle levende organismer er beslektet via felles avtamning, og derfor stammer de alle fra forfedre arter, som vandrer rundt i verden og diversifiserer, og genererer det fantastiske biologiske mangfoldet til organismer (Darwin, 1859). (20)
Mange typer organismer som lever i et miljø, "men med mindre du har en større kjent referansesekvens, er det veldig vanskelig å sette disse forskjellige tingene inn i en evolusjonært rammeverk (21)
 Koonin:
Koonin:
Argumenter for en LUCA som ikke kunne skilles fra en moderne prokaryotisk celle har blitt presentert, sammen med scenarier som viser LUCA som en mye mer primitiv enhet (Glansdorff, et al., 2008).
Vanskeligheten i problemet kan ikke overvurderes. Faktisk er alle kjente celler komplekse og forseggjorte. De enkleste kjente cellulære livsformene, de bakterielle (og de eneste kjente arkaiske) parasittene og symbiontene, utviklet seg tydelig ved nedbrytning av mer komplekse organismer; imidlertid har selv disse flere hundre gener som koder for komponentene i en fullverdig membran; replikerings-, transkripsjons- og oversettelsesmaskinerier; et komplekst celledelings-apparat; og i det minste noen sentrale metabolske veier. Som vi allerede har diskutert, er de enkleste frittlevende celler betydelig mer komplekse enn dette, med minst 1.300 gener.
Til tross for alle vanskeligheter og usikkerhetsfaktorer ved evolusjonære rekonstruksjoner, gir personanalyse kombinert med mindre formell innsats for rekonstruksjon av den dype fortiden til bestemte funksjonelle systemer, ingen alvorlig tvil om at LUCA allerede hadde minst flere hundre gener. I tillegg til de nevnte 'gylne 100' gener involvert i ekspresjon, består dette mangfoldige genkomplementet av mange metabolske enzymer, inkludert veier for den sentrale energi-metabolismen og biosyntese av aminosyrer, nukleotider og noen koenzymer, samt noen avgjørende membran-proteiner, slik som underenhetene til signalgjenkjennings-partikkelen (SRP) og H + - ATPase.
Fra og med 2014 sitter Koonin i den rådgivende redaksjonen for Trends in Genetics, og er medredaktør i en tilgjengelig åpen journal 'Biology Direct'. Han satt i redaksjonen for bioinformatikk fra 1999-2001. Koonin er også et rådgivende styremedlem i bioinformatikk ved 1000-fakultetet.
DNA-replikering over taxa, side 193: 2016
Genomsekvensering av celler fra livets tre domener, bakterier, arkeaer og eukaryoter, avslører at de fleste av kjernens replisomkomponenter utviklet seg to ganger uavhengig. Dermed deler ikke bakteriekjernens replisome-enzymer en felles stamfar med de analoge komponentene i eukaryoter og archaea, mens archaea og det eukaryotiske kjerne-replisom maskineriet deler en felles stamfar
Koonin, the logic of chance -side 331:
 Det rekonstruerte genrepertoaret til LUCA har også gapende hull. De to mest sjokkerende er
Det rekonstruerte genrepertoaret til LUCA har også gapende hull. De to mest sjokkerende er
(i) fraværet av nøkkelkomponentene i DNA-replikeringsmaskineriet, nemlig polymerasene som er ansvarlige for initiering (primaser) og forlengelse av DNA-replikasjon og for gap-fylling etter fjerning av 'primer', og de viktigste DNA-helikasene (Leipe, et al., 1999), og
(ii) fravær av de fleste enzymer fra lipid-biosyntese. Disse essensielle proteinene klarer ikke å gjøre det til det rekonstruerte genrepertoaret til LUCA fordi de respektive prosessene i bakterier, på den ene siden, og archaea, på den annen side, katalyseres av forskjellige, ikke-relaterte enzymer og, i tilfelle av membranfosfolipider, gir kjemisk forskjellige membraner.
Bakterier og archaea har membraner laget av vannavvisende fettmolekyler. Enkle fettmolekyler har en tendens til å snu seg rundt, noe som gjør membranen lekk, slik at både bakterier og archaea festes på en vannelskende fosfatgruppe for å stabilisere molekylene og gjøre membranene deres ugjennomtrengelige. De tok imidlertid veldig forskjellige ruter. Bakterielle membraner er laget av fettsyrer bundet til fosfatgruppen, mens arkeiske membraner er laget av isoprener bundet til fosfat på en annen måte. Dette antyder at membranene deres utviklet seg uavhengig. Dette fører til noe paradoksalt: Hvis LUCA allerede hadde en ugjennomtrengelig membran for å utnytte proton-gradienter, hvorfor ville dens etterkommere uavhengig ha utviklet to forskjellige typer ugjennomtrengelig membraner? (17)
 Grassos kommentar: Sjokkerende og bemerkelsesverdig:
Grassos kommentar: Sjokkerende og bemerkelsesverdig:
DNA-replikeringsmaskineriet er viktig i alle domener, og det er også lipidbiosyntese for cellemembraner. Det er ikke mulig at de første cellene dukket opp uten membraner og DNA-replikasjon i en LUCA, og deretter utviklet seg forskjellige membraner og DNA-replikasjon, hver for seg.
Det betyr at de minst flere hundre gener som innehas i alle livets tredomener, måtte ha kommet fram på en konvergent måte (det vil si hver for seg, de ville ha blitt til med samme genom, proteom og metabolom bortsett fra lipid-biosyntese og DNA-replikasjon som var de to eneste distinkte delene som skilte seg fra de andre domenene. Dette er vanskelig å selge når man fremkaller evolusjon. Enda mer når bare ikke-styrte tilfeldige mekanismer var tilgjengelig: sjanse og hell. Hvis en celletypes fremdukking hadde vært veldig usannsynlig, forestill deg den samme skjedde tre ganger, hver for seg. Som Stephen J. Gould skrev i Wonderful Life: The Burgess Shale and the Nature of History:
"... Ingen finale kan spesifiseres i starten, ingen vil noensinne forekomme en gang til på samme måte, fordi enhver vei fortsetter gjennom tusenvis av usannsynlige stadier. Endre enhver tidlig begivenhet, aldri så lite og uten tilsynelatende betydning på den tiden, og evolusjonen utvikler seg i en helt annen kanal. (1)
Grassos kommentar -fortsetter: Verken en LUCA er troverdig, eller naturlig fremvoksende tre atskilte livets domener, ved delvis konvergent vis. Den eneste rasjonelle forklaringen er en designer som lager livets tre domener hver for seg, og bruker det samme verktøysettet der det er nødvendig, og et eget divergerende verktøysett for andre deler.
Bør det ikke være bevis på at en LUCA aldri har eksistert, og at livets tre domener måtte oppstå separat gjennom en intelligent designer?
Den utbredte rollen til ikke-enzymatiske reaksjoner i cellulær metabolisme
Sekvenser av glykolytiske enzymer er forskjellige mellom Archaea og Bacteria / Eukaryotes (14)
Membraner utgjør også en av de mest sta oppgavene i all celleutvikling. Kort tid etter oppdagelsen av Archaea ble det innsett at disse organismer skiller seg påfallende fra bakteriene i kjemien til deres membranlipider. Archaea lager plasmamembranene av isoprenoide underenheter, bundet av eterbindinger til glyserol-1-fosfat; derimot benytter bakterier og eukarya fettsyrer bundet av esterbindinger til glyserol-3-fosfat. Det er noen få unntak fra regelen. Archaea-membraner inneholder ofte fettsyrer, og noen dypt forgrenende bakterier, som Thermotoga, favoriserer isoprenoideter-lipider (men selv de kobler etrene til glyserol-3-fosfat). Dette mønsteret av lipid-sammensetning, som grupperer bakterier og Eukarya sammen på den ene siden og Archaea på den andre, står i skarp kontrast til det som kan forventes fra det universelle treet, som setter Eukarya med Archaea (18) LUCA halloo ....
----
Prokaryot evolusjon og livets tre er to forskjellige ting
Konseptet med et livets tre er utbredt i evolusjonær litteratur. Det stammer fra å forsøke å oppnå et storslått enhetlig naturlig system som gjenspeiler en tilbakevendende prosess med arter og avstamming for alle former for liv. Tradisjonelt opererer disiplinen til systematikk i et lignende hierarki av todelte (noen ganger multi-forgrente) kategorier. Antagelsen om et universelt livets tre er avhengig av at evolusjonsprosessen er trelignende gjennom alle livsformer og hele den biologisk tid. I prokaryoter gjør de det ikke. Prokaryot evolusjon og livets tre er to forskjellige ting, og vi må behandle dem som slike, i stedet for å ekstrapolere fra makroskopisk liv til prokaryoter. I det følgende vil vi vurdere denne omstendigheten fra filosofiske, vitenskapelige og epistemologiske perspektiver, og antar at fylogeni valgte en enkelt modell som en 'gjensitter' fra den moderne syntese av evolusjon.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2761302/
I eukaryoter består plasmamembranen av steroler og karbohydrater. I prokaryoter inneholder plasmamembranen ikke karbohydrater eller steroler. Prokarotiske membraner har bare noen få typer fosfolipider, mens eukaryote membraner har over 6 forskjellige fosfolipider, så vel som andre typer lipider. Prokaryote membraner har ofte ikke kolesterol inne i den hydrofobe kjernen, mens eukaryote membraner bruker koresterol for å regulere fluiditeten. Eukaryot cellemembran er i utgangspunktet trilamellar med dobbelt lag fosfolipid. Det er asymmetrisk. Den har iboende og ytre proteiner som også hjelper til med transport over membranen. Den har andre komponenter som kolesterol for å opprettholde fluiditet i membranen. Hvor som prokaryotisk eller bakteriell cellemembran er sammensatt av peptidoglycan som er tverrkjede av N acetyl-glykosamin og muraminsyre.
Opprinnelsen og den tidlige utviklingen av eukaryoter i lys av fylogenomikk
Opprinnelsen til eukaryoter er en enorm gåte og en stor utfordring for evolusjonær biologi. Det er et skarpt skille i den organisatoriske kompleksiteten til cellen mellom eukaryoter, som har kompleks intracellulær kompartementalisering, og til og med de mest sofistikerte prokaryoter (archaea og bakterier), som ikke gjør det. En typisk eukaryot celle er omtrent 1000 ganger større i volum enn en typisk bakterie eller arkea, og fungerer under forskjellige fysiske prinsipper: Fri diffusjon har liten rolle i eukaryote celler, men er avgjørende i prokaryoter [7, 8]. Oppdelingen av eukaryote celler støttes av et forseggjort endomembran-system og av det actin-tubulin-baserte cytoskjelettet. Det er ingen direkte motstykker til disse organellene i archaea eller bakterier. Det andre kjennetegnet ved den eukaryote cellen er tilstedeværelsen av mitokondrier, som har en sentral rolle i energitransformasjon og utfører mange tilleggsroller i eukaryote celler, som signalering og celledød.
https://genomebiology.biomedcentral.com/articles/10.1186/gb-2010-11-5-209
Livets nettverk: genomets begynnelse og evolusjon
Den raske veksten av genom-sekvensdata siden midten av 1990-tallet gir nå enestående detaljer på det genetiske grunnlaget for livet, og katalyserer ikke overraskende den mest grunnleggende revurderingen av opprinnelse og evolusjon siden Darwins tid. Flere artikler i dette temanummeret hevder at Darwins 'livets tre' nå er best sett på som en tilnærming - en ganske adekvat som en beskrivelse av noen deler av den levende verden (f.eks. Morfologisk komplekse eukaryoter), men mindre nyttig andre steder (f.eks. virus og mange prokaryoter ); Faktisk går én av forfatterne våre lenger og kunngjør 'dødsfallet' av Darwins tre som en hypotese om mangfoldet og tilsynelatende naturligheten til hierarkiske ordninger for grupper av levende organismer.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2874017/
Å rive opp livets tre
Charles Darwin hevdet for mer enn hundre år siden at alle moderne arter avvek fra et mer begrenset sett av forfedre grupper, som selv utviklet seg fra enda færre forfedre og så tilbake til begynnelsen av livet. I prinsippet kan altså forholdet mellom alle levende og utdøde organismer bli representert som et slektstre. De fleste moderne forskere er enige. Mange vil til og med hevde at de generelle trekkene til dette treet allerede er kjent, helt ned til roten - en ensom celle, kalt livets siste universelle felles forfader, som levde for omtrent 3,5 til 3,8 milliarder år siden. Konsensusvisningen kom ikke lett, men har blitt allment akseptert i mer enn et tiår. Likevel blåser det dårlig vind. Til alles overraskelse har funn gjort de siste årene begynt å kaste alvorlig tvil om noen aspekter av treet, spesielt på skildringen av forholdene nær roten.
http://labs.icb.ufmg.br/lbem/aulas/grad/evol/treeoflife-complexcells.pdf
 Nylig oppdagede 'Orphan gener' trosser evolusjon (2)
Nylig oppdagede 'Orphan gener' trosser evolusjon (2)
En viktig kategori av 'useriøse' genetiske data som fullstendig trosser evolusjonære spådommer er den vanlige forekomsten av taksonomisk begrensede gener, ellers kjent som 'foreldreløse gener'. Disse blir nå oppdaget i sekvenseringen av alle genomer.
Mange flercellede dyr deler lignende sett med gener som produserer proteiner som utfører relaterte biokjemiske funksjoner. Dette er et vanlig trekk ved målrettet konstruerte systemer. I tillegg til disse standard-genene, har alle organismer som hittil er testet, også unike sett med gener som er spesifikke for den typen skapning.
Forfatterne av en nylig gjennomgangs-artikkel, publisert i Trends in Genetics, om foreldreløse gener uttalte: "Sammenlignende genomanalyser indikerer at hver taksonomisk gruppe hittil studert, inneholder 10-20% av gener som mangler gjenkjennelige homologer [lignende kolleger] i andre arter. " (1)
Disse foreldreløse genene blir også funnet å være spesielt viktige for spesifikke biologiske tilpasninger som samsvarer med økologiske nisjer i forhold til skapningens interaksjon med omgivelsene.(2) Problemet for den evolusjonære modellen av animalsk opprinnelse er det faktum at disse DNA-sekvensene dukker opp plutselig og fullt funksjonelle, uten spor av evolusjonær forfedre (DNA-sekvensforløpere i andre tilsynelatende relaterte organismer). Og flere nye studier i både fisk og insektgenomer fremhever nå dette viktige faktum.
Hva er livets tre?
Et universelt livets tre (LT) har lenge vært et mål for molekylære fylogenetikere, men nettikulering på nivå av gener og muligens på nivåer av celler og arter gjør enhver enkel tolkning av en slik TOL, spesielt som brukt på prokaryoter, problematisk. (12) En av de mange måtene mikrobiologi setter den ny-darwinistiske syntesen i fare, er ved å true med å "oppheve livets tre (TOL)" [1]. Lateral genoverføring (LGT) er mye hyppigere enn de fleste biologer ville ha forestilt seg før for rundt 20 år siden, så fylogenetiske trær basert på sekvenser av forskjellige prokaryote gener er ofte forskjellige. Hvordan man driller ut fra slike motstridende data, blir noe som kan svare til et enkelt, universelt livets tre, problematisk. Dessuten, siden mange viktige evolusjonære overganger involverer slektsfusjoner på et eller annet nivå, er treets egnethet (et mønster av etterfølgende bifurkasjoner) som en oppsummering av livets historie usikker [2–4].
http://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1005912
Blekkspruter ‘er romvesener’, slutter forskere etter DNA-undersøkelse
http://metro.co.uk/2015/08/12/octopuses-are-aliens-scientists-decide-after-dna-study-5339123/
Ikke for å 'freake deg ut' eller noe, men forskere har nettopp avslørt at åtte-armede blekkspruter er så rare at de i utgangspunktet er romvesener.
Den første fullstendige genomssekvensen viser at åtte-armede blekkspruter (ikke tiarmet) er helt forskjellige fra alle andre dyr - og deres genom viser et slående nivå av kompleksitet med 33 000 proteinkodende gener identifisert, mer enn hos et menneske.
Der tenkte vi at det var freaky nok da de lærte å åpne syltetøyglass.
 Amerikansk forsker Dr Clifton Ragsdale, fra University of Chicago, sa:; Blekkspruten ser ut til å være helt forskjellig fra alle andre dyr, til og med andre bløtdyr, med sine åtte prehensile armer, sin store hjerne og sine smarte problemløsende metoder.
Amerikansk forsker Dr Clifton Ragsdale, fra University of Chicago, sa:; Blekkspruten ser ut til å være helt forskjellig fra alle andre dyr, til og med andre bløtdyr, med sine åtte prehensile armer, sin store hjerne og sine smarte problemløsende metoder.
‘Den avdøde britiske zoologen Martin Wells sa blekkspruten er en fremmed. I denne forstand beskriver altså papiret vårt det første sekvenserte genomet fra en alien. ’
Blekksprut: Hva er de?
De bor i hvert hav på nesten alle dyp og har en rekke funksjoner som minner om sci-fi-romvesener.
Disse inkluderer sugekledde gripe-tentakler, svært mobile, kameralignende øyne som er følsomme for polarisert lys, sofistikerte kamuflasjesystemer som endrer hudfarge og mønstre, jetfremdrift, tre hjerter og muligheten til å regenerere skadde lemmer.
Forskerne anslår at to-delte blekksprut genomet inneholder 2,7 milliarder basepar - de kjemiske enhetene til DNA - med lange strekninger av gjentatte sekvenser.
 Dekoding av genomet til en alien
Dekoding av genomet til en alien
https://www.eurekalert.org/pub_releases/2015-08/oios-dtg081215.php
Foruten gjenkjennelige gener, består store deler av genomet av regulatoriske nettverk som styrer hvordan gener uttrykkes i celler. I blekkspruten ble nesten halvparten av genomet funnet å være sammensatt av mobile elementer kalt transposoner, en av de høyeste andelene i dyreriket. Transposoner replikerer og beveger seg rundt for seg selv, forstyrrer eller forbedrer genuttrykk og letter omorganisering av genorden. Forskerne fant at mange av dem var spesielt aktive i blekksprutens nervesystem. "Hox" -genene, involvert i embryonal utvikling hos alle dyr, er et spesielt dramatisk eksempel. Selv om de er samlet sammen i de fleste dyr, inkludert andre bløtdyr, er de spredt i utklipp i blekkspruten, noe som antagelig muliggjør utviklingen av den allsidige blekksprut-kroppsplanen.
Antagelig. Ja. Eller med andre ord, gjettverk som alltid ... Arkitekturen til en kroppsplan må være helt fra begynnelsen. Alt virker, eller ingenting virker. Spørsmålet er, hvor kommer informasjonen til denne omorganiseringen av gener fra? Etter mitt syn er den eneste rasjonelle forklaringen forsettlig (intelligent) design.
Er Rotifers gen-tyver eller unikt konstruert? (1)
Verktøyene for DNA-sekvensering blir billigere å bruke og mer produktive enn noensinne, og flommen av DNA-sammenligningsresultater mellom organismer som kommer, blir en hengemyr for det evolusjonære paradigmet. For å støtte det, tyr biologer til stadig mer absurde forklaringer på avvik. Et godt eksempel på denne lureriet er i et nylig DNA-sekvenseringsprosjekt utført i et mikroskopisk akvatisk flercelledyr kalt en rotifer. (1)
I dette arbeidet målrettet forskerne mot gensekvensene som uttrykkes som proteiner for DNA-sekvensering fordi genomet var for stort og komplekst til å sekvensere og montere alt DNA. De registrerte over 61.000 gensekvenser som ble uttrykt fra rotorer dyrket under stressede og ikke-stressede forhold. Av disse kunne de bare finne sekvenslikheter mellom rotifer og andre skapninger for 28 922 sekvenser (mindre enn halvparten). Forskerne kastet de ukjente DNA-sekvensene ut av analysen siden de ikke-lignende genene var nye, tilsynelatende spesifikke for rotifer, og i det vesentlige vanskelige for evolusjon å forklare.
 Av de 28 922 sekvensene som de kunne oppnå samsvar i en offentlig database med andre skapningers DNA- og proteinsekvenser, passet ikke en betydelig andel (mer enn i noen annen skapning sekvensert) evolusjonære forventninger av vanlig avstamning. Ytterligere komplisert dette bildet ble rotifer-gensekvensene funnet i et mangfoldig antall ikke-rotifer-skapninger! Noen av skapningene som hadde genmatch til rotifer inkluderte en rekke planter, andre flercellede dyr, protister (komplekse encellede dyr), arkeaer, bakterier og sopp. Evolusjonister har to muligheter for å kategorisere disse uvanlige gen-sammenfall basert på deres naturalistiske forutsetninger. For det første kan de si at disse genene utviklet seg uavhengig i separate skapninger i en hypotetisk prosess kalt 'konvergent evolusjon'. I tilfeller der det bokstavelig talt er hundrevis av disse DNA-sekvensene som dukker opp i flere organismer, blir dette scenariet så lite sannsynlig, at selv evolusjonister har for store problemer med å forestille seg det. Det andre alternativet kalles 'Horisontal GenOverføring', eller HGO. Dette innebærer overføring av gener, kanskje via en slags mikrobiell verts-organisme, lik en bakterie. (2)
Av de 28 922 sekvensene som de kunne oppnå samsvar i en offentlig database med andre skapningers DNA- og proteinsekvenser, passet ikke en betydelig andel (mer enn i noen annen skapning sekvensert) evolusjonære forventninger av vanlig avstamning. Ytterligere komplisert dette bildet ble rotifer-gensekvensene funnet i et mangfoldig antall ikke-rotifer-skapninger! Noen av skapningene som hadde genmatch til rotifer inkluderte en rekke planter, andre flercellede dyr, protister (komplekse encellede dyr), arkeaer, bakterier og sopp. Evolusjonister har to muligheter for å kategorisere disse uvanlige gen-sammenfall basert på deres naturalistiske forutsetninger. For det første kan de si at disse genene utviklet seg uavhengig i separate skapninger i en hypotetisk prosess kalt 'konvergent evolusjon'. I tilfeller der det bokstavelig talt er hundrevis av disse DNA-sekvensene som dukker opp i flere organismer, blir dette scenariet så lite sannsynlig, at selv evolusjonister har for store problemer med å forestille seg det. Det andre alternativet kalles 'Horisontal GenOverføring', eller HGO. Dette innebærer overføring av gener, kanskje via en slags mikrobiell verts-organisme, lik en bakterie. (2)
Hvilken fallitt mot felles avstamning fra en hovedstrøms forsker:
Den biologiske Big Bang-modellen for de store overgangene i evolusjonen
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1973067/
Store overganger i biologisk evolusjon viser det samme mønsteret med plutselig fremvekst av forskjellige former på et nytt nivå av kompleksitet. Forholdet mellom hovedgrupper i en ny klasse av biologiske enheter er vanskelig å tyde og ser ikke ut til å passe til det tremønsteret som, etter Darwins opprinnelige forslag, fortsatt er den dominerende beskrivelsen av biologisk evolusjon. De aktuelle tilfellene inkluderer opprinnelsen til komplekse RNA-molekyler og proteinfold; store grupper av virus; archaea og bakterier, og de viktigste linjene innenfor hvert av disse prokaryote domenene; eukaryote supergrupper; og dyre-rekkene. I hver av disse sentrale forbindelsesledd (nexuses) i livets historie ser de viktigste 'typene' ut til å vises raskt og fullt utstyrt med signatur-egenskapene til det respektive nye nivået av biologisk organisasjon. Ingen mellomliggende 'karakterer' eller mellomformer mellom forskjellige typer kan påvises. Vanligvis tilskrives dette mønsteret kladogenese komprimert i tid, kombinert med den uunngåelige erosjonen av det fylogenetiske signalet.
Argumentet til det nedveltede evolusjonstreet
1. Den grunnleggende prinsippet i evolusjonsteorien er at arter utviklet seg i henhold til det evolusjonære treet; den ene etter den andre utviklet seg som et geneologisk slektstre.
2. Siden Darwin har imidlertid vitenskapen fortsatt å dokumentere unntak og avvik - arter som ikke passer pent inn i evolusjonsmønsteret.
- For eksempel, arter som i mange henseender ser ut til å være ganske like, som evolusjonister har plassert på nabokvister av det evolusjonære treet, blir rutinemessig funnet å ha dype forskjeller. Her er et eksempel:
a. I 2010 utgav en artikkel i tidsskriftet Nature [4] resultatene av en DNA-sjimpansestudie med implikasjoner som var veldig overraskende for det vitenskapelige samfunnet fordi resultatet av forskningen motsa den lenge holdte hypotesen om deres likhet.
b. Allerede tittelen oppsummerte forskningsfunnene: "Sjimpanse og menneskelige Y-kromosomer er bemerkelsesverdig divergerende i struktur og geninnhold."
c. Sjimpanse-DNA-sekvensen for et kromosom ble samlet og orientert basert på et Y-kromosom-kart/rammeverk bygget for sjimpanse og ikke menneskelig. Som et resultat kunne sjimpanse-DNA-sekvensen da bli mer nøyaktig sammenlignet med det menneskelige Y-kromosomet.
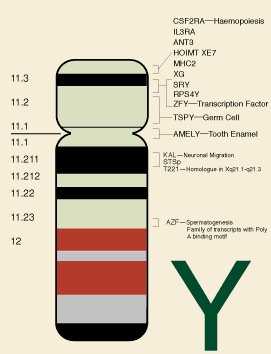 d. Sjimpansen og humane Y-kromosomer hadde en dramatisk forskjell i geninnhold på 53 prosent. Med andre ord manglet sjimpansen omtrent halvparten av genene som ble funnet på et humant Y-kromosom.
d. Sjimpansen og humane Y-kromosomer hadde en dramatisk forskjell i geninnhold på 53 prosent. Med andre ord manglet sjimpansen omtrent halvparten av genene som ble funnet på et humant Y-kromosom.
c. Forskerne forsøkte også å avgjøre om det var noen forskjell i faktiske genkategorier, og de fant en sjokkerende 33 prosent forskjell.
e. Det menneskelige Y-kromosomet inneholder en tredjedel flere genkategorier - helt forskjellige klasser av gener - sammenlignet med sjimpanser.
f. Fordi praktisk talt alle strukturelle aspekter av menneskekromosomene og sjimpanse Y var forskjellige, var det vanskelig å komme til et samlet likhetsestimat mellom de to. Forskerne postulerte en samlet 70 prosent likhet, som ikke tok hensyn til størrelsesforskjeller eller strukturelle forskjeller. Dette ble gjort ved å konkludere med at bare 70 prosent av sjimpansesekvensen kunne være justert med den menneskelige sekvensen - uten å ta hensyn til forskjeller innenfor linjene. I.O.W. 70 prosent var et forsiktig estimat, spesielt når man vurderer at 50 prosent av de menneskelige gener manglet fra sjimpansen, og at regionene som hadde en viss likhet var lokalisert i helt andre mønstre. Når alle aspekter av ikke-likhet - sekvenskategorier, gener, genfamilier og genposisjon - tas i betraktning, er det trygt å si at den generelle likheten var lavere enn 70 prosent.
g. I Nature-artikkelen kan vi lese, "Faktisk, ved 6 millioner års separasjon, er forskjellen i MSY-geninnhold i sjimpanse og menneske mer sammenlignbar med forskjellen i autosomalt geninnhold i kylling og menneske, etter 310 millioner års separasjon."
h. Så det menneskelige Y-kromosomet ser like forskjellig ut fra en sjimpanse som de andre menneskelige kromosomene gjør fra en kylling. Og for å forklare hvor alle disse forskjellene mellom mennesker og sjimpanser kom fra, er troende på en evolusjonsfortelling i det store og hele, tvunget til å finne på historier om store kromosomale omlegginger og rask generering av store mengder mange nye gener, sammen med tilhørende regulatorisk DNA.
i) Men siden hvert respektive Y-kromosom ser ut til å være fullt integrert og gjensidig avhengig av sin vertsorganisme, er den mest logiske slutningen av Y-kromosomdataene at mennesker og sjimpanser hver for seg ble spesielt laget som forskjellige skapninger.
- På den annen side blir arter som åpenbart er ganske forskjellige, som evolusjonister har plassert på fjerne grener av det evolusjonære treet, ofte funnet å ha dype likheter.
a) Mennesker, Arabidopsis (En slekt av sennepsfamilien som har hvite, gule eller lilla blomster) og nematoder (usegmenterte ormer med langstrakt avrundet kropp spisset i begge ender) har alle omtrent samme antall gener.
b) Et forskerteam fra Heidelberg fra European Molecular Biology Laboratory [EMBL] [1], sammenlignet menneske- og bananflue-introner [2] med de fra en rundorm som antas å være 600 millioner år gamle. Overraskende nok var introner allerede i ormene fra begynnelsen av deres framtreden, og forble de samme helt til den menneskelige linjen, og endret seg raskt og mistet mange av dem bare i andre arter som insekter. En av forskerne bemerket: "Nå har vi direkte bevis for at gener allerede var ganske komplekse i de første dyrene, og mange hvirvelløse dyr har redusert en del av denne kompleksiteten." Og en annen sa: "Ormens gener er veldig lik menneskelige gener ... Det er et mye annerledes bilde enn vi har sett fra de raskt utviklende artene som har blitt studert så langt." I tillegg har genomet også blitt "bevart i løpet av de siste en halv milliard år." I sin forskning forklarte de ikke hvordan de tidlige kambriske rundormene fikk sin kompleksitet og evne til å forbli uendret i millioner av år. Oppdagelsen endrer åpenbart evolusjonstreet.
c) Molekylære evolusjonstrær passer ofte ikke til et morfologi-basert evolusjonstre. For eksempel er det flere TRAF-gener hos mennesker og Drosophila, og en åpenbar spådom av Darwins modell er at det må være et forfedregen i en felles forfedre organisme som de moderne TRAF-gener ble hentet fra. I virkeligheten passer imidlertid ikke et TRAF-gen fra Hydra til kriteriene til et forfedren, som må være noe av en blanding av alle humane TRAF-familier, men hører heller tydelig til hovedgruppen av TRAF-gener sammen med human TRAF1, TRAF2, TRAF3 og TRAF 5, mens human TRAF4 og spesielt TRAF6 tilhører forskjellige grupper sammen med Drosophila TRAFs. [3]
3. I mange år forsøkte evolusjonister å forklare den voksende listen over motsetninger ved hjelp av deres evolusjonære tremodell. Men det er åpenbart at dette var en øvelse i å tvinge bevisene til å passe teorien i stedet for omvendt.
4. I løpet av de siste årene har evolusjonister endelig begynt å nedtone vekten av deres ikoniske evolusjonære tremodell. Det som imidlertid ikke endres, er deres insistering på at evolusjon er et faktum.
 5. Dermed læres studentene også i dag at arten faller inn i det forventede tremønsteret. Men noen vågale forfattere begynner å nevne denne umulige, forbudte arkeologien.
5. Dermed læres studentene også i dag at arten faller inn i det forventede tremønsteret. Men noen vågale forfattere begynner å nevne denne umulige, forbudte arkeologien.
6. For få år siden rapporterte for eksempel Telegraph at «Charles Darwins livets tre er 'feil og villedende'.
- De mener begrepet villeder oss fordi hans [Darwins] teori begrenser og til og med tilslører studiet av organismer og deres forfedre. ...
- Forskere sier at selv om biologien i store deler av de siste 150 årene i stor grad har vært opptatt av å fylle ut detaljene på treet, er det nå foreldet og må kastes. ...
- "I lang tid var den hellige gralen å bygge et livets tre. Vi har overhodet ingen bevis for at livets tre er en realitet".
- Mer fundamentalt antyder nyere forskning at evolusjonen av dyr og planter ikke akkurat er trelignende heller. ...
- Dr. Rose sa: "Livets tre begraves høflig - det vet vi alle. Det som er mindre akseptert er at hele vårt grunnleggende syn på biologi må endres." Han sier at biologi er langt mer kompleks enn vi trodde, og å møte denne kompleksiteten vil være like skummelt som de konseptuelle omveltningene fysikerne måtte ta ombord tidlig på 1900-tallet.
7. Motstridende bevis ble/blir kontinuerlig diskuterert åpent. Men ingen av dem tillates å kaste tvil om selve evolusjonsteorien. Som artikkelen rapporterte:
8. "Hvis du ikke har et livets tre, hva betyr det for evolusjonær biologi? Først er det veldig skummelt - men de siste par årene har folk begynt å frigjøre seg." Både han og medforsker Dr Ford Doolittle understreket at nedgradering av livets tre ikke betyr at evolusjonsteorien er feil, bare at evolusjon ikke er så ryddig som vi vil tro.
9. Teorien må modifiseres og forstørres gjentatte ganger for å prøve å tilpasses dataene. På et tidspunkt blir teorien lite mer enn en tautologi. Uansett hvilken oppdagelse som gjøres i biologien, må evolusjonen ha skapt den, uansett hvor motstridende og usannsynlig.
10. Imidlertid er slik tautologi en av feilene i logikken. Per definisjon:
"Tautologi i formell logikk refererer til en uttalelse som må være sant i enhver tolkning, ved selve konstruksjonen. I retorisk logikk er det et argument som bruker sirkulært resonnement, noe som betyr at konklusjonen også er dens eget premiss. Typisk er forutsetningen ganske enkelt omformulert i konklusjonen, uten å legge til ytterligere informasjon eller avklaring. Strukturen til slike argumenter er A = B, derfor A = B, selv om forutsetningen og konklusjonen kan formuleres annerledes, slik at den ikke umiddelbart er tydelig som sådan."
11. Dermed er den eneste logiske forklaringen på forskjeller mellom lignende arter og likheter mellom forskjellige arter, en involvering av en intelligent designer, som bruker lignende genetiske mønstre. Denne kaller alle mennesker Gud.
12. Gud eksisterer.
Referanser:
Notes:
1. Vertebrate-type intron-rich genes in the marine annelid Platynereis dumerilii F. Raible, K. Tessmar-Raible, K. Osoegawa, P. Wincker, C. Jubin, G. Balavoine, D. Ferrier, V. Benes, P. de Jong, J. Weissenbach, P. Bork and D. Arendt.
2. intron - Part of a gene whose sequence is transcribed but not present in a mature mRNA after splicing.
3. Mali B, Frank U. Hydroid TNF-receptor-associated factor (TRAF) and its splice variant: a role in development.Mol Immunol. (2004) 41:377-84
4. Hughes, J.F. et al. 2010. Chimpanzee and human Y chromosomes are remarkably divergent in structure gene content. Nature. 463 (7280): 536-539.
Omtrent samtidig konkluderte evolusjonsbiologen Dalhousie University, W. Ford Doolittle, at lateral genoverføring mellom gamle organismer betydde at molekylær fylogeni kanskje aldri kunne oppdage det "sanne treet" i livet, ikke fordi det bruker feil metoder eller feil gener, "men fordi livets historie ikke riktig kan vises som et tre." Han konkluderte: "Kanskje det ville være lettere, og på sikt mer produktivt, å forlate forsøket på å tvinge" molekylære data "inn i formen som ble gitt av Darwin." I stedet for et tre 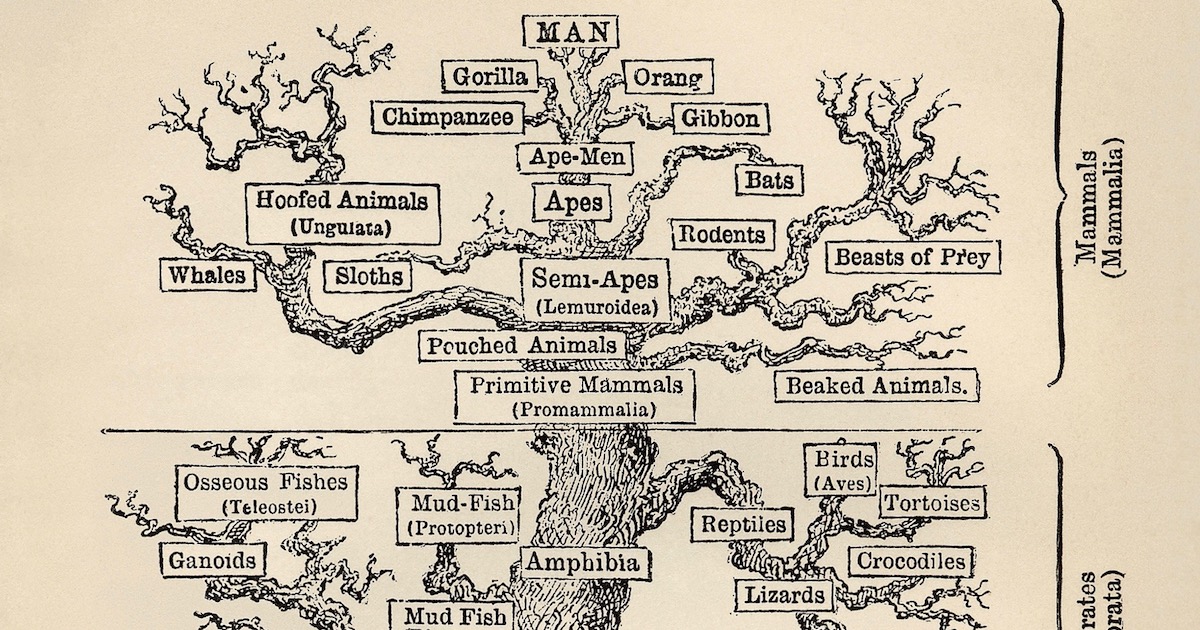 foreslo Doolittle "et nett- eller nettlignende mønster." (10)
foreslo Doolittle "et nett- eller nettlignende mønster." (10)
Kontroversen om livets universelle tre fortsetter. I 2002 foreslo Woese at biologi skulle gå utover Darwins doktrine for felles avstamning. I 2004 skrev han: "Roten til det universelle treet er en gjenstand som er et resultat av å tvinge evolusjonskurset inn i en trerepresentasjon når denne representasjonen er upassende." I 2004 foreslo Doolittle og hans kolleger å erstatte livets tre med en nettlignende 'syntese av livet', og i 2005 anbefalte de at "andre representasjoner enn et tre skulle undersøkes." I mellomtiden fortsetter andre forskere å forsvare hypotesen om at den universelle stamfaren eksisterte, men var kompleks snarere enn enkel (11)
DR ROSE SA: "LIVETREET BLIR GJORT POLITISK - VI VET ALLE DET. HVA SOM ER MINDRE AKSEPTERTT ER AT HELE VÅRT GRUNNLEGGENDE SYN PÅ BIOLOGI ER I BEHOV FOR Å ENDRES". Han sier at biologi er mer komplisert enn vi trodde og å møte dette kompliserte vil være like skummelt som de konseptuelle opphevelser som fysikere måtte ta ombord i det tidlige 20. århundre.
Roten til det universelle livets tre er ikke pålitelig (5)
Flere sammensatte universelle trær som er koblet sammen med en stamfar gendublisering, har blitt brukt til å forankre livets universelle tre. I alle tilfeller viste denne roten seg å være i den eubakterielle gren. Gyldigheten av resultatene fra komparativ sekvensanalyse har imidlertid nylig blitt stilt spørsmålstegn ved, spesielt når det gjelder eldgamle fylogenier. For eksempel har det blitt vist at flere eukaryote grupper er feilplassert i ribosomalt RNA eller forlengelses-faktortrær på grunn av ulik evolusjonshastighet og mutasjons-metning.  Videre har tillegg av nye sekvenser til datasett, ofte gjort tilsynelatende rimelige fylogenier forvirrende. Vi har dermed revidert alle sammensatte proteintrær som har blitt brukt til å rote det universelle livets tre til nå (forlengelsesfaktorer, ATPaser, tRNA-syntetaser, karbamoylfosfat-syntetaser, signalgjenkjenningspartikkelproteiner) med oppdaterte datasett. Generelt var de to prokaryote domenene ikke monofyletiske med flere avvikende grupperinger på forskjellige nivåer av treet. Videre motsatte de respektive fylogeniene seg hverandre, slik at ulike ad hoc-scenarier (paralogi eller lateral genoverføring) må foreslås for å oppnå det tradisjonelle søsterskap Archaebacteria - Eukaryota. Enda viktigere er at alle markørene er sterkt mettet med hensyn til aminosyre-substitusjoner. Ettersom fylogenier utledet fra mettede datasett, er ekstremt følsomme for forskjeller i evolusjonære rater, kan nåværende fylogenier som brukes til å forankre det universelle livets tre være forutinntatt av fenomenet lang tiltrekning. Siden den eubakterielle grenen alltid var den lengste, kunne eubakteriell forankring forklares med en tiltrekning mellom denne grenen og den lange grenen av utgruppen. Til slutt foreslo vi at en eukaryotisk forankring kunne være en mer fruktbar arbeidshypotese, da den for eksempel gir en enkel forklaring på den høye genetiske likheten til Archeabakteria og Eubakteria, utledet fra fullstendig genomanalyse.
Videre har tillegg av nye sekvenser til datasett, ofte gjort tilsynelatende rimelige fylogenier forvirrende. Vi har dermed revidert alle sammensatte proteintrær som har blitt brukt til å rote det universelle livets tre til nå (forlengelsesfaktorer, ATPaser, tRNA-syntetaser, karbamoylfosfat-syntetaser, signalgjenkjenningspartikkelproteiner) med oppdaterte datasett. Generelt var de to prokaryote domenene ikke monofyletiske med flere avvikende grupperinger på forskjellige nivåer av treet. Videre motsatte de respektive fylogeniene seg hverandre, slik at ulike ad hoc-scenarier (paralogi eller lateral genoverføring) må foreslås for å oppnå det tradisjonelle søsterskap Archaebacteria - Eukaryota. Enda viktigere er at alle markørene er sterkt mettet med hensyn til aminosyre-substitusjoner. Ettersom fylogenier utledet fra mettede datasett, er ekstremt følsomme for forskjeller i evolusjonære rater, kan nåværende fylogenier som brukes til å forankre det universelle livets tre være forutinntatt av fenomenet lang tiltrekning. Siden den eubakterielle grenen alltid var den lengste, kunne eubakteriell forankring forklares med en tiltrekning mellom denne grenen og den lange grenen av utgruppen. Til slutt foreslo vi at en eukaryotisk forankring kunne være en mer fruktbar arbeidshypotese, da den for eksempel gir en enkel forklaring på den høye genetiske likheten til Archeabakteria og Eubakteria, utledet fra fullstendig genomanalyse.
Tidlig evolusjon uten livets tre (6)
Det er mer med evolusjon enn det som passer til noe tre. For å forstå store overganger i tidlig evolusjon, trenger vi kanskje ikke et livets tre i det hele tatt. Men vi må holde ideene våre testbare med data fra genomer eller andre uavhengige data for å holde nesen festet til slipesteinen til observasjoner. Den veldig tidlige evolusjonen av livet er for det meste skrevet på kjemispråket, hvorav noen (uten tvil) fremdeles fungerer i dag i moderne metabolisme hvis vi ser på de riktige gruppene. Miljøene og utgangsmaterialet som jorden hadde å tilby for å drive tidlig kjemi, er variabler som bare geokjemikere med rimelighet kan begrense. Man kan anføre at acetogener (clostridial firmicutes) og hydrogenotrofe metanogener (euryarkareoter) huser de forfedre tilstandene til mikrobiell fysiologi i henholdsvis eubakterier og arkea-bakterier, og noen trær er kompatible med den oppfatningen, og det er også fordelingen av primitive energibesparende mekanismer. . Men gitt en overgang fra elementene på tidlig jord til replikerende celler, ser ikke utviklingen av prokaryoter ut til å spille seg ut langs grenene til et fylogenetisk tre. For eksempel undersøkte Whitman biologien og mangfoldet av prokaryoter, og viste et rRNA-tre for å diskutere spørsmål om klassifisering; men forgrenings-rekkefølge i det treet spiller ingen rolle i hans diskusjon om mangfold eller underliggende evolusjonære prosesser. Hvis det er retningen vi er på vei i, er det ikke dårlig. Men å ha eukaryoter sittende på en gren i livets rRNA-tre i stedet for på to, slik de burde være (eller tre når det gjelder planter med plastider), er langt nok utenfor merket om at vi burde streve for et bedre representasjon av forholdet mellom eukaryoter og de to typer prokaryoter som de stammer fra.
Eukaryoter er genetiske kimærer, og rollen som mitokondrier i opprinnelsen til den kimærismen er tydelig. Eukaryoter er komplekse, og mitokondriens sentrale rolle i begynnelsen av den kompleksiteten (i motsetning til en sentral rolle fagocytose) virker stadig vanskeligere å bestride av energi- grunner. Det etterlater lite rimelig alternativ til synspunktet om at verten for mitokondriens opprinnelse var en prokaryot, i det enkleste av konkurrerende alternativer en archea-bakterium. Antikken til anaerob energimetabolisme og sulfidmetabolisme blant eukaryoter passer godt sammen med nyere utsikt over proterozoisk havkjemi. En utfordring gjenstår i datanettverk av genomer som inkluderer lateral genoverføring mellom prokaryoter og opprinnelsen til eukaryoter i samme graf. Å spore tidlig evolusjon uten livets tre gir mye mer frihet til å utforske ideer enn å tenke med et tre for hånde. Ideene må likevel generere spådommer og være testbare, ellers er de ikke vitenskap. Hvis vi sjekker tankene våre for raskt mot et tre, hvis sannhet ingen kan bestemme uansett, begynner treet å bestemme hvilke tanker vi kan ha eller ikke, og hvilke ord vi kan eller ikke kan bruke. Skal et livets tre politi styre tankene våre? Å jobbe uten ett er et alternativ.
Mønsterpluralisme og livets tre-hypotese (7)
Darwin hevdet at et unikt inkluderende hierarkisk mønster av forhold mellom alle organismer basert på deres likheter og forskjeller [Livets tre (LT)] var et naturfaktum, for hvilket evolusjon, og spesielt en forgreningsprosess med nedstigning og modifikasjon, var forklaringen. Imidlertid er det ingen uavhengige bevis for at den naturlige ordenen er  et inkluderende hierarki, og inkorporering av prokaryoter i LT er spesielt problematisk. De eneste datasettene vi kan konstruere et universelt hierarki fra, inkludert prokaryoter, gener av sekvenser, er ofte uenige og kan sjelden bevises å være enige. Hierarkisk struktur kan alltid pålegges eller ekstraheres fra slike datasett, ved hjelp av algoritmer designet for å gjøre det, men i sin base hviler det universelle LT på en uprøvd antagelse om mønster som, gitt det vi vet om prosessen, er usannsynlig å være hovedsakelig sant. Dette er ikke for å si at likheter og forskjeller mellom organismer ikke skal redegjøres for av evolusjonære mekanismer, men nedstigning med modifisering er bare én av disse mekanismene, og et enkelt trelignende mønster er ikke det nødvendige (eller forventede) resultatet av deres kollektiv drift. Mønsterpluralisme (erkjennelsen av at forskjellige evolusjonære modeller og representasjoner av relasjoner vil være hensiktsmessige, og sanne, for forskjellige rekker eller på forskjellige nivåer eller for forskjellige formål) er et attraktivt alternativ til den virkelighetsfjerne jakten på ett eneste ekte Livets Tre (LT).
et inkluderende hierarki, og inkorporering av prokaryoter i LT er spesielt problematisk. De eneste datasettene vi kan konstruere et universelt hierarki fra, inkludert prokaryoter, gener av sekvenser, er ofte uenige og kan sjelden bevises å være enige. Hierarkisk struktur kan alltid pålegges eller ekstraheres fra slike datasett, ved hjelp av algoritmer designet for å gjøre det, men i sin base hviler det universelle LT på en uprøvd antagelse om mønster som, gitt det vi vet om prosessen, er usannsynlig å være hovedsakelig sant. Dette er ikke for å si at likheter og forskjeller mellom organismer ikke skal redegjøres for av evolusjonære mekanismer, men nedstigning med modifisering er bare én av disse mekanismene, og et enkelt trelignende mønster er ikke det nødvendige (eller forventede) resultatet av deres kollektiv drift. Mønsterpluralisme (erkjennelsen av at forskjellige evolusjonære modeller og representasjoner av relasjoner vil være hensiktsmessige, og sanne, for forskjellige rekker eller på forskjellige nivåer eller for forskjellige formål) er et attraktivt alternativ til den virkelighetsfjerne jakten på ett eneste ekte Livets Tre (LT).
En vanlig evolusjonær opprinnelse for funksjonelle moduler av hale-bakteriofag og bakterielle maskiner(8)
Den dramatiske divergensen av bakteriofag-genomer er en hindring som ofte forhindrer påvisning av homologi mellom proteiner og dermed bestemmelse av fylogenetiske koblinger mellom bakteriofagene. For eksempel er sekvenslikhet mellom Siphoviridae hoved-hale proteiner, som har blitt demonstrert eksperimentelt for å danne bakteriofagens-halerør, ofte ikke observerbart.
Fage Proteomic Tree: en genombasert taksonomi for fager (9)
Bakteriofager inneholder ikke en ribosomal sekvens som gjør at de kan plasseres på livets universelle tre, og til dags dato har de ikke hatt fordel av sitt eget genbaserte taksonomiske system. Tidligere forsøk på å klassifisere og måle fagmangfold basert på genetiske markører har opplevd begrenset suksess. Selv om strukturelle proteiner (f.eks. kapsider) hypotetisk kunne tjene som grunnlag for fagtaksonomi (27, 29, 41, 60), er de svært forskjellige og inneholder, i motsetning til rDNA, ikke konserverte regioner som gjør at de lett kan identifiseres. Dette begrenser nytten av disse proteinene som markører for biologisk mangfoldstudier. Vi viser her at det ikke finnes noe enkelt gen i alle fager som kan brukes som grunnlag for et klassifiseringssystem.
Hva eksakt viser genetisk likhet? (1)
Opprinnelse til gener med uløst avstamning
Hovedproblemet: Uintelligent vs. intelligent mekanisme
 Mitt håp er at en dag å tenkning om Darwinistisk teori vil bli tydeligere på det offentlige torget. Husk at Darwin kom med to påstander: (1) alle levende vesener stammer fra en eller noen få opprinnelige forfedre, og (2) mekanismen som driver endringene blant arter er den blinde, ikke-styrte mekanismen for naturlig utvalg.
Mitt håp er at en dag å tenkning om Darwinistisk teori vil bli tydeligere på det offentlige torget. Husk at Darwin kom med to påstander: (1) alle levende vesener stammer fra en eller noen få opprinnelige forfedre, og (2) mekanismen som driver endringene blant arter er den blinde, ikke-styrte mekanismen for naturlig utvalg.
Den kontroversielle påstanden er selvfølgelig den andre - ideen om at en rent materiell mekanisme, uten intelligens involvert, er ansvarlig for all den genetiske informasjonen som er nødvendig for livet (DNA) og dermed for hele livets mangfold.
Sekvenslikhet alene beviser IKKE felles stamfedre
98,8% DNA-sekvenslikheten mellom sjimpanser og mennesker som Clines-referanser ikke engang etablerer krav ett (felles stamfedre). Og "du trenger ikke å ta mitt ord for det," som LeVar Burton alltid pleide å si om Reading Rainbow.
Bilde: Livets tre -veltet (?)
Som Francis Collins, leder av prosjektet som kartla det menneskelige genomet, har skrevet om DNA-sekvenslikheter: "Dette beviset alene beviser selvfølgelig ikke en felles stamfar" fordi en intelligent sak kan gjenbruke vellykkede designprinsipper".
Vi vet dette fordi vi er intelligente agenter selv, og vi gjør dette hele tiden. Vi tar instruksjoner vi har skrevet om en ting og bruker dem til en annen. Likheten er ikke resultatet av en blind mekanisme, men snarere resultatet av vår intelligente aktivitet.
Noen designforkjempere synes bevisene for felles stamfedre er gode (f.eks. Michael Behe), mens andre - med henvisning til fossilregistertet, spesielt The Cambrian Explosion - ikke gjør det. Men ingen av gruppene mener at sekvenslikhet alene beviser verken felles stamfar eller den darwinistiske mekanismen, slik mange vitenskapelige forfattere i vår tid virker ivrige etter å formode.
Vanlige DNA-sekvenser: Bevis for evolusjon eller effektiv design?
Med fremveksten av moderne bioteknologi har forskere vært i stand til å bestemme den faktiske sekvensen til de omtrent tre milliardene DNA-basene (A, T, C, G) som utgjør det menneskelige genomet. De har også sekvensert genomene til mange andre typer skapninger. Forskere har prøvd å bruke disse nye DNA-dataene for å finne likheter i DNA-sekvensene til skapninger som angivelig er relatert gjennom evolusjonær avstamning, men gir genetiske likheter bevis for evolusjon?
Oversettelse og bilder ved Asbjørn E. Lund